觸覺與視覺比起來,好像不那麼重要,因為我們要辨認物體似乎多靠視覺。但是請你想想以下幾個情境,也許你會改變這個想法。
想像你在暗室中摸到一籃水果,你是不是可以不靠視覺,單純藉著觸覺知道哪個是香蕉,哪個是蘋果?甚至從表面的光滑度、堅硬度及特有的形狀知道這是哪一類的頻果?答案當然是肯定的。再想像當你伸出手要去抓眼前看到的物體時,結果卻抓不到東西,亦即沒有任何觸覺上的訊息回饋,此時你會相信觸覺?還是相信視覺?當然,幾乎沒有例外的,大家多會相信觸覺。可見觸覺比視覺更能取得我們本身的信賴,也許『seeing is believing』應該改成『feeling is believing』了。
觸覺研究的先驅者David Katz(1925)很早便指出,要靠觸覺去感受某一表面的質地時,手指與物體表面之間必須發生摩擦移動。蓋因皮膚上含有數以千計的機械感受器(mechanoreceptors),不同的受器負責不同的觸感,有的負責粗糙度的感覺,有的負責硬度的感覺,不一而論。因此接下來我們將分別討論這些受器的生理機制與心理物理特性。
觸覺的心理物理特性
觸覺敏感度的測量最早是由Max von Frey在1986年提出,沿用至今。他發現不同粗細、不同長度的毛髮需要不同的力矩才恰好可以彎曲,根據這些毛髮的力距特性,預先篩選出不同力距的毛髮製成類似探針的儀器,然後在各處皮膚上做探測。如此可以知道不同的皮膚位置原來有不同的觸感力道,我們稱這種靜態的觸感為觸覺敏感度(tactile sensitivity)。有趣的是,我們發現嘴唇的觸覺敏感度優於背部,而且這種敏感度有性別差異,即女性優於男性。
後來的研究者(Verillo,1962;Van Doren,Pelli,and Verillo,1987)將Max von Frey的儀器修改成可以產生不同振觸頻率的刺激(vibrotactile stimuli),結果發現皮膚最高的敏感觸頻可達200Hz。這種觸頻敏感度的測量甚至可以成為臨床上診斷中毒者的週邊神經系統受傷害程度。
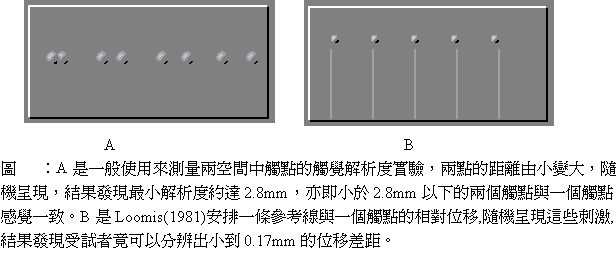
還有一種稱為兩點覺閾(two-point threshold)的測量,最早由Weinstein(1968)進行,結果發現指尖可以區分達2mm的刺激間距,手臂最好則是30mm,到了背部就變成70mm了,這類兩點觸覺位置區分能力稱為觸感敏銳度(touch acuity)。兩點覺閾實驗結果顯示,指尖的觸覺受器應該是密而小的,反之,手掌上的受器就比較疏而大(Vallbo & Johansson,1978;Kandel & Jessell,1991)。
直覺上可以推想在兩點觸感敏銳度高的皮膚上應該有很好的刺激定位能力(localization ability),事實上的確如此。不過,對於單點刺激的定位能力可能比兩點觸感敏銳度更好,這可以由下列實驗作業表現出來:Jack Loomis(1981)設計如圖 的實驗材料與程序,發現有參考線時的單一觸點位置之偏移量的偵測比單純兩點觸覺解析度要好,這個結果類似視覺的hyperacuity現象。
雖然觸覺刺激的空間定位能力極佳,但是仍然有位置上的錯覺發生!最有名的例子是Aristotle's illusion,即將食指、中指交叉之後,以一隻筆幹同時碰觸兩指端,此時會感覺到似乎有兩隻筆幹碰觸的感覺(相對於兩指未交叉時的筆觸感覺,則更加明顯)。Benedetti(1985)認為這種觸錯覺意味著觸覺有類似視覺的拓樸對應機制存在,即相鄰的觸覺感受對應相鄰的皮質區,不相鄰的觸覺感受則送到不相鄰兩區。正如同一物體投影在兩眼相對應區時,我們會覺得看見一個物體,反之,兩眼相對應區投影兩個不同物體時,我們將會產生雙重影像。
再觀察當你矇住眼睛,由另一個人以鉛筆鈍端在你背部或手心寫字時,你很難判斷出他寫什麼字。除非他寫字時從頭到尾筆端都沒離開皮膚,一筆畫完成,此時就比較容易判斷他在畫什麼圖案了。如果他每劃一點就將筆端離開皮膚一下,再畫下一點,那麼你幾乎不可能『認出』他畫的圖案,可見依賴觸覺的型態辨認具有時效性。
再來觀察我們對於表面質地的觸感特性。Susan J.Lederman(1981)…………….
觸覺生理基礎
因為觸覺主要靠手,而且手掌觸覺受器有相當大的大腦相對應處理區,所以我們將把觸覺生理的介紹聚焦在手上的觸覺受器與相關大腦活動。
基本上我們可以依照解剖位置,把手掌的觸覺神經分成兩束,一束稱為Ulnar神經束,其神經軸突源自於手掌下方包括小指及半個無名指,另一束稱Median神經束,軸突源自於拇指、食指、中指與半個無名指。像視覺細胞一樣,這些神經每一個都有其獨特的感覺接受域及其反應特性。Vallbo與Hagbarth(1968)首先在人體上直接記錄手部觸覺受器的神經衝動訊號,並依照其反應的時間特性與空間特性,分別將這些神經細胞做了以下分類:
就時間特性而言可以分兩類,一類是慢適應型(slowly adapting fibers,以下簡稱SA),亦即當壓力刺激呈現時,它就開始反應,只要刺激一直存在,它就一直保持大約一致的神經反應,直到刺激消失;另一類是快速適應型(rapidly adapting fibers,以下簡稱RA),亦即只當刺激出現一剎那時會出現反應,並在刺激移除那一剎那,再出現一次強烈反應。
就空間特性而言,也可分兩類,一類是侷限型(punctate fibers),即該觸覺受器的接受域較小,並有清楚的約成卵圓形的邊界;另一類是擴散型(diffuse fibers),與侷限型相反,它沒有清楚的接受域的邊界。因此兩類時間特性與兩類空間特性交叉混合成四類觸覺神經:SA侷限型(SAI)、RA侷限型(RAI)、SA擴散型(SA II)與RA擴散型(PC)。其中,RA侷限型最接近皮膚表層,其受器表面還有膜囊包圍,被稱為Meissner Corpuscles;再深入皮膚時則是SA侷限型受器,結構上無膜囊包住,稱為Merkel Disks;再深入一層則是SA擴散型受器,結構上也無膜囊,被稱為Ruffini Endings;而最內層的則是RA擴散型受器,有大大的膜囊包住,被稱為Pacinian Corpuscles。
Johannsonn與Vallbo(1979)進一步測定主觀觸感與這些觸覺神經的關係發現,當輕輕觸碰手掌時,只有RA型神經的反應被記錄到。如果刺激再強一些,再持續一些時間,則SA型的神經反應才會出現。而且SA型神經的反應可以約略反應主觀觸壓強度,但並不是完全相關。如果直接刺激SA型神經,則受試者會報告有一股輕軟的觸壓持續在手掌皮膚上;直接刺激RA型神經則會引發皮膚上有震盪的觸壓感。我們將這些主觀感受與觸覺受器的生理特性總整理如下表:
空間特性 時間 特性與感受 |
侷限型(R.F.小) |
擴散型(R.F.大) |
慢速適應型 最佳反應頻率 觸感 |
Merkel Disks(SAI) 0.3-3Hz 慢壓 |
Ruffini Endings(SA II) 15-400Hz 皮膚拉扯 |
快速適應型 最佳反應頻率 觸感 |
Meissner Corpuscles(RAI) 3-40Hz 輕觸 |
Pacinian Corpuscles(PC) >500Hz 快速震動 |
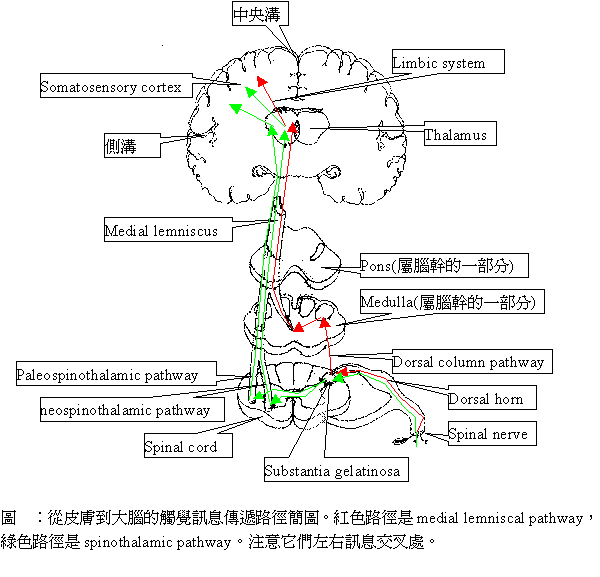
這四類受器的神經衝動所傳達的第一站是脊髓,透過脊髓的背根(dorsal root),神經衝動可以向上傳到大腦,也可以從腹根(ventral root)引發運動神經元產生肌肉抽動的反射動作。本章並不打算細談脊髓反射弧,而將重點放在上傳大腦的途徑上。基本上,上傳路徑又分為二條,一條是由脊髓傳到腦幹(brainstem),再到medial lemniscus,中途在延髓(medulla)處發生左右訊息交叉,再上達parietal lobe中的somatosensaory cortex,稱為medial lemniscal pathway(或稱dorsal column),是由比較粗大、有髓鞘的(myelinated)、傳導速度快的Aβ型脊髓神經纖維攜帶觸覺訊息與肢體位置訊息(proprioception),也是本章的重點;另一條稱為spinothalamic pathway,是負責攜帶溫度與疼痛訊息的,又可分為paleospinothalamic(paleo-是舊的意思)與neospinothalamic (neo-是新的意思)兩條路徑,舊的路徑是由比較細小、無髓鞘、傳導速度慢的C型纖維負責,新的路徑主要是由細小的、有髓鞘的、傳導慢的Aδ型纖維負責。重要的是新舊兩條路徑都一起到達thalamus及邊緣系統(Limbic system),再上傳至somatosensory皮質區。但是關於這方面的研究很少,此處還沒有足夠的知識談它,因此這裡也將忽略這些討論。
從thalamic nuclei傳出的神經,會投射到初級somatosensaory區(以後簡稱S1),也會投射一部份到二級somatosensaory區(以後簡稱S2),不過S2的訊息大部分仍是來自S1的神經投射。基本上S1可分出兩類區域,一類包括area 3b與area 1接受來自皮膚表層侷限型受器的訊息,另一類包括area 2與area 3a,是接受來自皮膚深層擴散型受器的訊息,而且它們的接受域特性十分複雜。
科學家對觸覺皮質細胞之接受域研究最多的都在猴子身上。他們發現猴子thalamus對於手臂之觸覺接受域有on-center-off-surround的分佈特性,類似於視覺系統中的LGN的細胞反應(Mountcastle & Powell,1959)。如果在皮質部(somatosensory cortex)做電生理記錄,則不只有center-surround的接受域分佈特性,還有對不同方位邊界敏感的特性,這類似於視覺上的簡單細胞與複雜細胞的反應特性,還有一些皮質細胞只對於物體碰觸指尖時的特定運動方向(例如由左滑到右;或由右滑到左)敏感(Hyvarinin & Poranen,1978)。這些觸覺細胞的反應特性顯示,視覺與觸覺兩類感覺訊息處理的方式可能存在共同的運作法則。
Wilder Penfield與Theodore Rasmussen(1950)這兩位神經外科醫生,恰好有機會對癲癇症患者做開腦摘除腫瘤手術時,被允許在病人大腦內做電刺激的反應記錄(這類記錄有利於手術後確認病人的其他腦部位功能是否受到手術影響,因此這類電刺激處理也是必要的)。結果發現,若將這些皮質區域所負責的肢體感覺對應區畫出來,幾乎成了一個腦中小人,只是這個小人的各個部位比例非常奇怪。如果你仔細看圖 ,應該會發現手指與臉、唇幾乎是放大倍率最高的,這似乎與該部位的功能有些關連。中國的針灸急救方中,對於中風病人的急救就是在十隻手指尖上(稱十宣穴)針刺放血,可能也與這些放大對應有些關連吧。
Craig(1985)則關心個體的預期是否影響觸覺辨認能力。他進行了以下的實驗。首先他設計了一個11mm x 27mm的方塊板,其中插了6x18根可由電腦控制的安全探針,令受試者將同一隻手的食指與中指同時輕放於上(見下圖,是由上而下的上視圖),探針的凹凸可以表現出不同字母形狀,實驗是要求受試者報告食指或中指探針的觸覺形狀。結果發現,當實驗者預先告訴受試者要注意哪隻指頭時,正確認出該指探針形狀的比率顯著高於未事先告知的情境。可見預期所產生的注意力會顯著提高指觸辨認能力。
有趣的是,當用不同手的指頭來做這個實驗時(即左手食指與右手食指,或反過來亦可),就不會發現上述由預期產生的效果。這似乎意味著左右手的處理可以同時分別由右腦與左腦進行,各腦半球有各自的注意力資源,不會互相影響。這類實驗結果對於視障、聽障的特殊教育教學上有極大啟發。